Rosalie Lawrence1, and Nick Kaplinsky1§
1Swarthmore College
§Correspondence to: Nick Kaplinsky (nkaplin1@swarthmore.edu)
Abstract
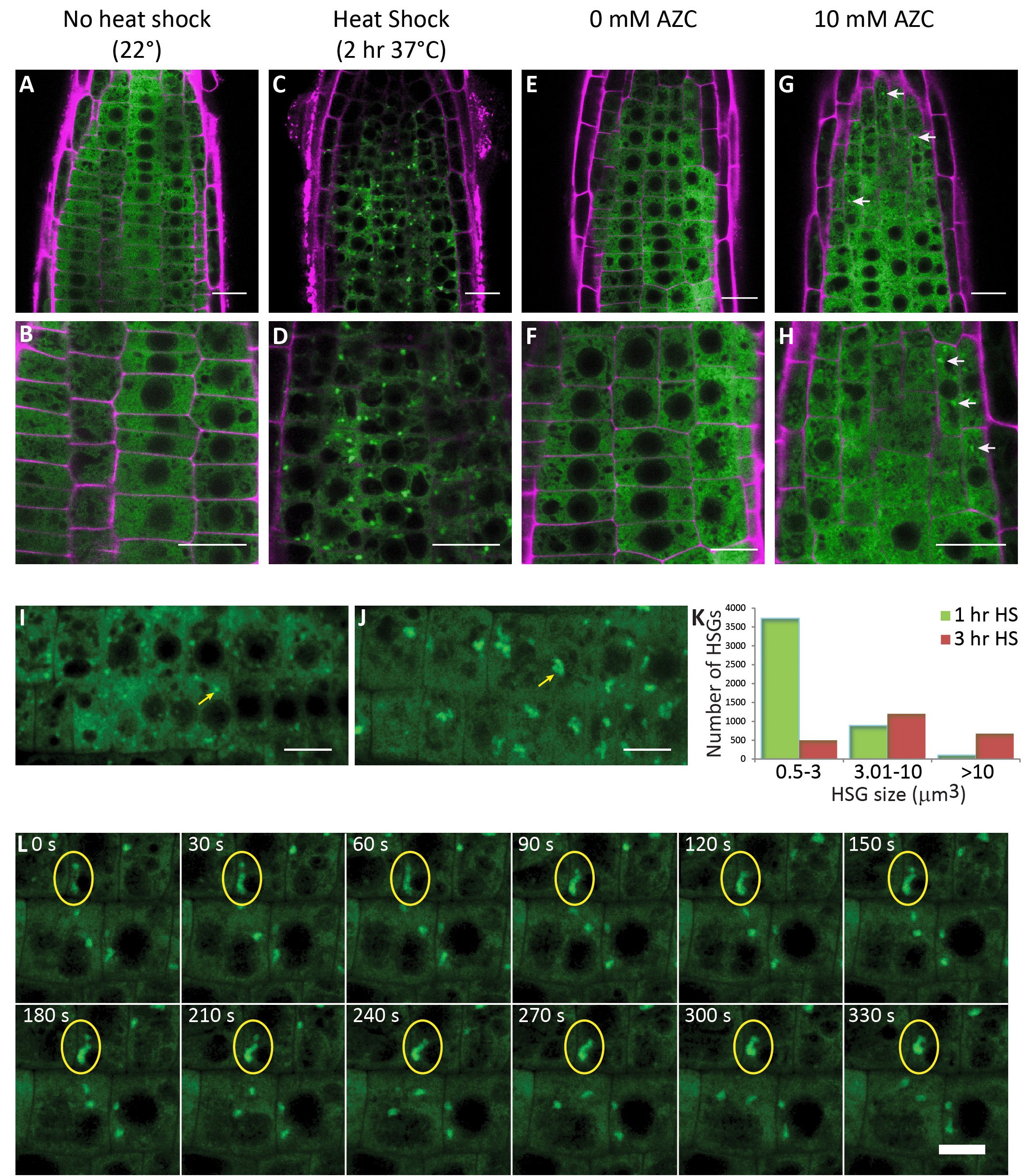
Description
Methods
Reagents
Extended Data
- Description: Maximal Z and Y projections of a BOB1:GFP time lapse movie with frames obtained every 30 seconds for a total of 50 minutes. The time lapse displays root cells between 144 and 194 minutes of HS treatment at 37oC. Yellow ovals indicate two granules that fuse over the course of this movie. Scale bar 10μm. . Resource Type: Audiovisual. DOI: 10.22002/D1.1457
Funding
Author Contributions
- Rosalie Lawrence: Conceptualization, Investigation, Methodology, Writing - review and editing, Writing - original draft
- Nick Kaplinsky: Conceptualization, Supervision, Writing - review and editing, Writing - original draft
Reviewed By
Anonymous
History
- Received: 5/31/2020
- Revision Received: 7/26/2020
- Accepted: 7/28/2020
- Published: 7/29/2020
Copyright
© 2020 by the authors. This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International (CC BY 4.0) License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation
PubMed Central: PMC7396158
PubMed: 32760882
microPublication Biology is published by
1200 E. California Blvd. MC 1-43 Pasadena, CA 91125
The microPublication project is supported by

The National Institute of Health -- Grant #: 1U01LM012672-01
microPublication Biology:ISSN: 2578-9430