Mackenzi Oswald1, Heino Hulsey-Vincent1, and Caroline (Lina) Dahlberg 1§
1Department of Biology, Western Washington University, Bellingham, WA, 98225, USA
§Correspondence to: Caroline (Lina) Dahlberg (lina.dahlberg@wwu.edu)
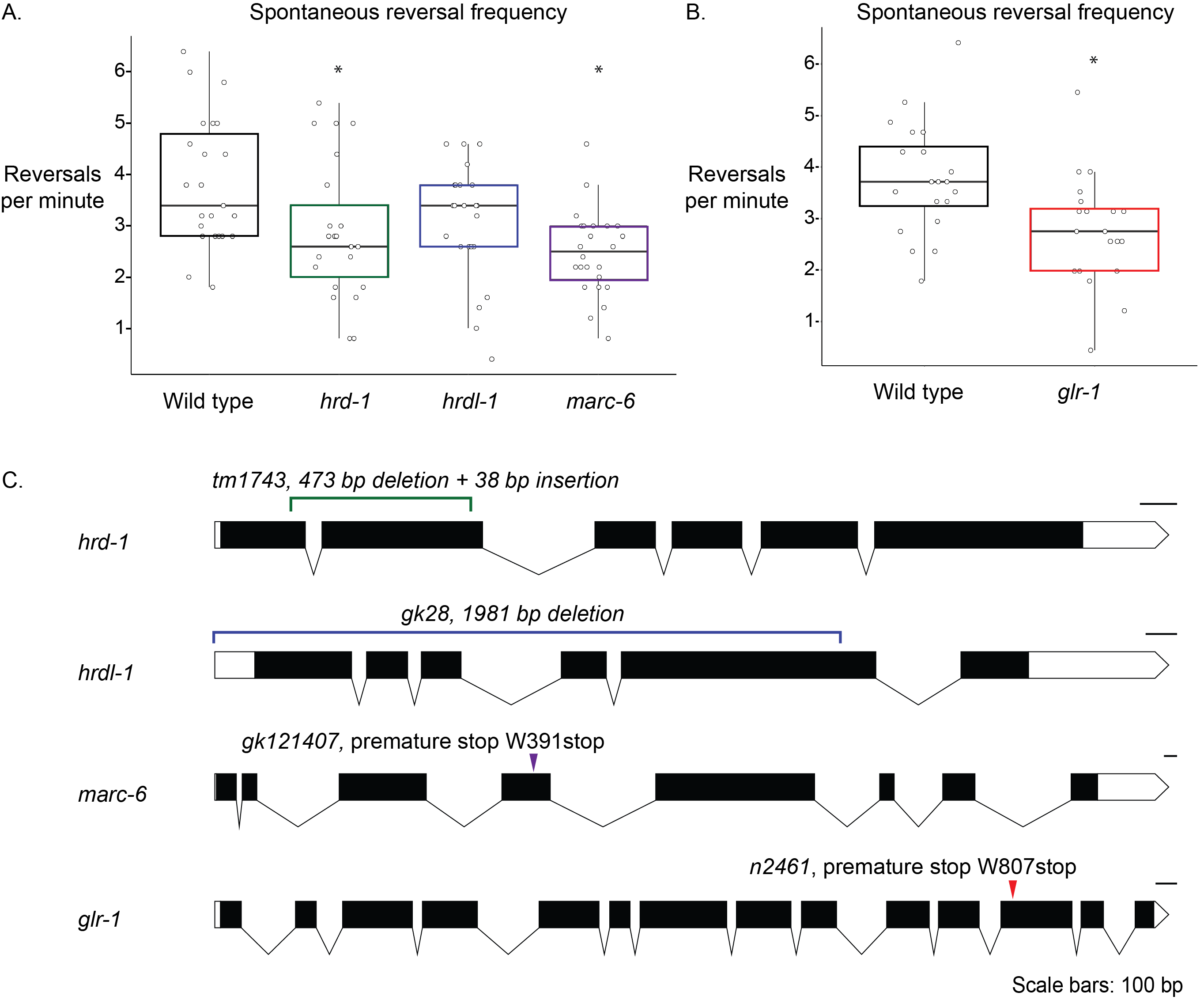
Description
Methods
Reagents
Acknowledgements
Funding
Author Contributions
- Mackenzi Oswald: Formal analysis, Funding acquisition, Investigation, Methodology, Writing - original draft, Writing - review and editing
- Heino Hulsey-Vincent: Formal analysis, Visualization, Writing - review and editing
- Caroline (Lina) Dahlberg : Conceptualization, Formal analysis, Funding acquisition, Methodology, Project administration, Resources, Supervision, Writing - review and editing
Reviewed By
Eric Luth, Anonymous
Nomenclature Validated By
Anonymous
Database Reference ID: WBPaper00060631
History
- Received: 6/2/2020
- Revision Received: 11/9/2020
- Accepted: 11/10/2020
- Published: 11/19/2020
Copyright
© 2020 by the authors. This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 International (CC BY 4.0) License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation
PubMed Central: PMC7704257
PubMed: 33274327
microPublication Biology is published by
1200 E. California Blvd. MC 1-43 Pasadena, CA 91125
The microPublication project is supported by

The National Institute of Health -- Grant #: 1U01LM012672-01